
by Thomas Fernandes

Last time, we explored how insects navigate and hunt using motion-based vision, and how these perceptual interfaces can themselves be exploited by dragonflies using motion camouflage to trick conspecifics.
In this essay, we shift from insects to spiders, and from motion cues to the many sensory strategies spiders use to sense their environment and guide their behavior. These developments culminate in the Portia spider, which has evolved one of the most remarkable visual systems known, along with cognitive abilities to match it. Harland and Jackson assembled three representative hunting scenarios involving Portia, drawn from repeated observations, which I will use as illustrative examples during the essay, while we try to understand their perceptual space.
Spiders are an unlikely place to look for visual sophistication. The first spiders evolved more than 300 million years ago, long before flying insects. Web-building spiders, which dominated early spider evolution, rely only minimally on vision. Instead, they externalize perception into silk. The web is not just a trap; it is an extended sensory organ. Web-dwelling spiders using aerial structures developed about 150 million years ago targeting flying insects.
In web-dwelling spiders, vibration replaces vision as the primary channel of information. Abiotic sources such as wind tend to displace large portions of the web at once in low frequency. Biotic sources like struggling prey, potential mates, invading conspecifics or even predators, originate from specific locations usually at higher frequencies.
The web transmits and amplifies all signals generated, while the spider “listens” with its legs, reconstructing a three-dimensional map of tension and displacement. It senses these displacements using slit sensilla, specialized strain sensors located at the joints of its eight legs, capable of detecting movements as small as 0.1 μm.
Spiders localize prey by comparing vibration patterns across their eight legs, which function as spatially distributed sensors. By analyzing differences in radial and transverse vibrations, the spider can estimate both the direction and distance of the source. Direction is inferred from the leg experiencing the greatest radial tension in the silk. Transverse vibrations, measured by comparing displacements across legs positioned at roughly 90° to each other, provide distance cues. Like a taut fabric pressed at one point, the deformation is steep and uneven nearby but shallow and uniform at a distance. The closer the prey, the sharper these cross-leg differences become.
This may seem complex to explain or model, but it is directly integrated as sensation for the spider and elicits almost instantaneous responses when a fly crashes into the web (video link). If the spider is waiting elsewhere, it must first return to the hub to accurately pinpoint the prey’s position.
This web already provides a rich perceptual space one that supports surprisingly complex behavior. During spinning, spiders adjust silk properties in different parts of the web and the geometry of the web itself is tuned to maximize information transfer to the central hub, where the spider typically waits. It does not wait passively but can actively probe the web.
Observing how spiders interpret weaker signals of insect struggling in the web (and not crashing) reveals that depending on the amplitude of the vibrations, spiders will either close in on the prey or interrogate the web by actively vibrating it to amplify signals before deciding to move.
Some species even construct decoys as shown here or in this video. Long before spiders became active hunters, they were already operating within an extended sensory world.
Today, not all spiders rely on webs; in fact, about half of known

spider species do not use webs for capture but are instead active hunters. Wandering spiders such as Cupiennius salei hunt without capture silk and are often nocturnal. Their vision is poor, but they make use of another vibration sensing organ of spiders, trichobothria. These are extremely fine hairs that detect air currents and vibrations directly. In general, wandering spiders have more trichobothria than web spiders for whom the airflow-sense is less important behaviorally. Two modalities dominate their perception.
The first is substrate vibration. Vibrations travel not only through a web but also through plants or bark. Cupiennius can signal a potential mate by generating vibrations that propagate along banana leaves and stems over several meters. These signals use frequencies low enough to travel with minimal attenuation through the plant, yet remain distinct from background noise.
The second is airflow. As an insect flies, it generates a smooth pressure field in front of it and a turbulent wake behind. As an insect approaches, Cupiennius senses the vibrations and orients toward the gentle front flow but does not strike. Only when the irregular wake passes over its hairs does it leap. The wake’s frequency structure reliably signals “living prey” rather than wind while also indicating the precise location of the fly: straight above. When deprived of its airflow sensors, Cupiennius never jumps into the air to catch prey, but it readily does so with its eyes covered. When the air flow is too strong the spider goes from a hunting response to a hiding response, sensing a bigger predator.
This solution works. It is robust, energetically efficient, and well suited to nocturnal life. But it has limits. Airflow and vibration are excellent for detecting that something is there. They are poor for identifying what it is doing, where it is oriented, or how it might respond next. The detection radius is also limited to about 25 cm.
Jumping spiders took a different path. They abandoned both webs and reliance on airflow cues and became active, visually guided hunters. Since visual acuity scales with eye size, small eyes should see poorly. Yet, 10 mm long jumping spiders routinely discriminate prey types, detect fine postural details, and plan precise jumps over many body lengths. What would have prompted such an evolution?
It was once proposed that high-acuity vision in jumping spiders evolved specifically for invading other spiders’ webs and eating first the insects caught in them and then the residing spiders themselves. A web-invading spider is at a massive disadvantage, because signal transmission is designed to carry information toward the hub and every web is structured differently, according to the preference of the residing spider.
Vision, by contrast, provides stable information about prey identity, location, and behavior, independent of web structure. Later phylogenetic work weakened the strongest version of this hypothesis, since high-acuity vision also evolved in lineages that do not invade webs. Still, among spider-eating lineages, increasing refinements in vision tend to accompany increasingly flexible predatory strategies. Here is an account from a spider with some of the most elaborate hunting sequences, Portia, illustrating how it exploits visual cues for web invasion.
“Portia sits near the edge of an orb web, looking across the sticky spirals toward the web’s architect, Gasteracantha sp., sitting at the hub. Gasteracantha is a distinctive spider. It is large, powerful, and has long horny spines on its abdomen that make it difficult for Portia to hold on to. Gasteracantha’s eyesight is too poor to recognize Portia as a predator. However, Gasteracantha has an acute ability to detect and interpret web signals. Portia’s task is to get within attacking distance without eliciting the wrong response from Gasteracantha. This large spider is fully capable of preying on Portia should it get the upper hand. Just walking across the web will not work for Portia. The resulting web signals will give it away.
So, Portia moves slowly onto the edge of the web, reaches out with its forelegs, and begins to pluck on the silk; but Gasteracantha does not move. Portia continues to make signals, but varies them. It plucks with different legs, plucks with its palps, varies the speed and the amplitude at which its appendages move, and it shakes the web by vibrating its abdomen up and down. Complex patterns are made by simultaneously moving different sets of appendages, with different appendages moving in different ways. By using any combination of its eight legs, two palps, and abdomen, Portia is capable of generating an almost unlimited repertoire of web signals. Eventually a signal may cause a reaction in Gasteracantha, and it may approach Portia. If the approach is not too fast, Portia will continue to signal, slowly drawing the prey spider closer. Portia avoids making web signals that elicit a fast approach because, when moving quickly, Gasteracantha is dangerous and more likely to become predator than prey. Luring Gasteracantha is a slow process, and close to an hour has already passed.
Then something happens to speed things up. A light tropical breeze gently rocks the web. The wind-induced web movements mask any fainter movements caused by Portia and the spider takes advantage of the smoke screen by stepping rapidly across the web toward Gasteracantha. This time, however, when the breeze dies down, Portia is still several centimeters from its prey.
Now Portia creates a smoke screen of its own by violently and repeatedly flexing all of its legs at the same time, the spider shakes the web much as the breeze did. Cloaked by a succession of such diversions, Portia closes the remaining distance. However, when it is about 3 mm from Gasteracantha, something goes amiss. Gasteracantha suddenly turns on Portia, lunging forward and grabbing one of Portia’s legs with its chelicerae (spider fangs). Portia leaps off the web, leaving the leg behind. After landing on the forest floor half a meter below, Portia looks up at the web and then climbs back to it. Once there, it repeats the entire process and this time succeeds in lunging at Gasteracantha. Portia quickly punctures Gasteracantha’s cuticle (external exoskeleton) with its fangs (injecting venom) and then lets go. Gasteracantha runs to the edge of the web and drops to the ground in an attempt to escape, but paralysis soon sets in. Portia drops to the ground on a dragline, walks in the direction of Gasteracantha, and scans the forest floor for the specific kind of spider it just attacked. Portia will bypass other potential prey placed in its path, continuing to search for the expected prey, in this case, Gasteracantha”- source
Jumping spider vision is among the most studied and here is what we know so far. Of their eight eyes, six secondary eyes are arranged around the head, together covering nearly 360 degrees. These monochromatic eyes are specialized for motion detection, guiding body turns and aligning the principal eyes with identified targets.
Functionally, these eyes actually resemble hearing in humans. A sudden sound triggers a turn of the head for investigation; a moving object triggers a turn of the spider. These eyes are not optimized for fine identification but for detecting where something is happening. Already this is a departure from most other spider that would rely on air vibrations. Jumping spider still have trichobothria providing them with “spider sense” detecting airflow near them. But vision allows for much longer detection radius in prey or predators.
The true marvel of vision resides in their two principal eyes. Across animals, visual acuity scales strongly with eye size. Falcons are exceptional among vertebrates, but relative to its size, a jumping spider’s visual acuity is even more extraordinary. There are three aspects of their unique vision.
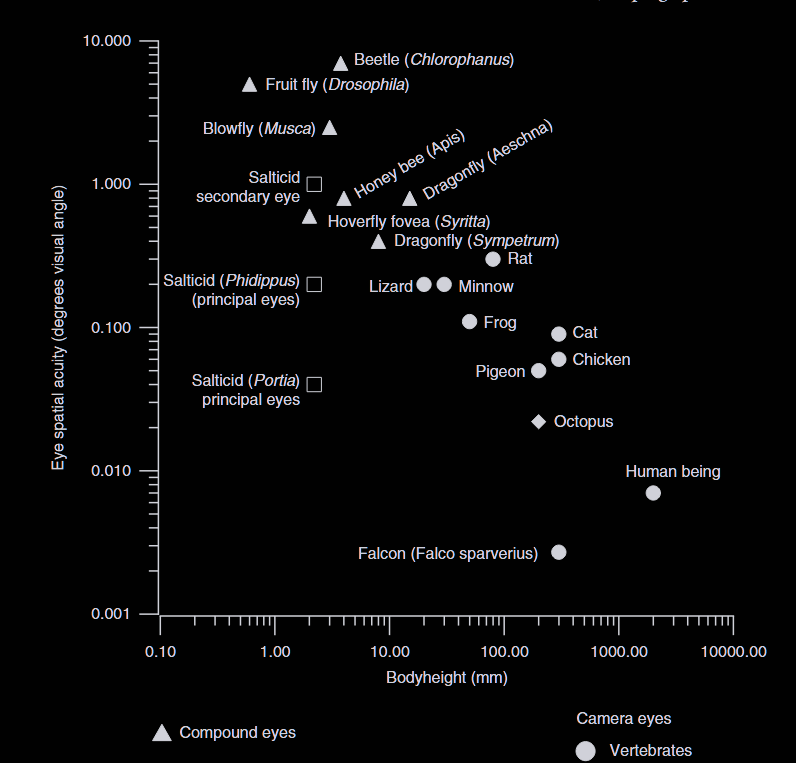
The first is magnification, which allows Portia to see tiny details with extraordinary precision. To project different features onto separate receptors, the spider couldn’t enlarge the image due to its small retina, so it uses magnification instead. Magnification trades a wide field of view for fine resolution, much like a telescope. The principal eyes reduce their visual field to about 6°, roughly the width of an espresso cup held at arm’s length. Within that tiny window however, magnification boosts resolution about thirtyfold.
To explore this narrow slice of the world, the retinas themselves move: muscles sweep them side-to-side and up-and-down, scanning the scene inside the fixed eye tubes. In effect, the spider carefully reads its environment line by line, building a detailed mental map. This slow, deliberate strategy shows why jumping spiders are not reactive hunters but patient stalkers, observing and planning before making a move.
The second is chromatic aberration. Lenses focus different wavelengths of light at slightly different distances. In large eyes, this difference is negligible, but in the tiny eyes of jumping spiders, it can introduce significant blur.
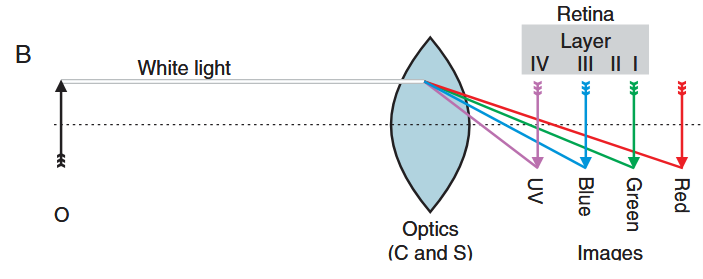
Rather than eliminating this problem, jumping spiders exploit it. The principal eye retina is layered, with photoreceptor layers positioned at different depths to match the focal planes of different wavelengths. Ultraviolet and blue are captured in the first layer, and green in the last two layers (layers are numbered from back to front). This arrangement requires precise organization within and across layers, allowing the spider to extract information from one layer while preserving light for the others.
Looking at Figure 5 you can notice that layer II in particular receives a consistently defocused image. What looks like a design flaw turns out to be a computational resource. By comparing the sharpness of images across both green layers, the spider can extract information about distance. The amount of defocus becomes a cue for absolute depth. Figure 6 highlights how distance can be computed by comparing images across the two layers. In most animals, defocus contributes only weakly to relative depth. Jumping spiders are the first known animals where it is used as a primary metric with impressive precision.
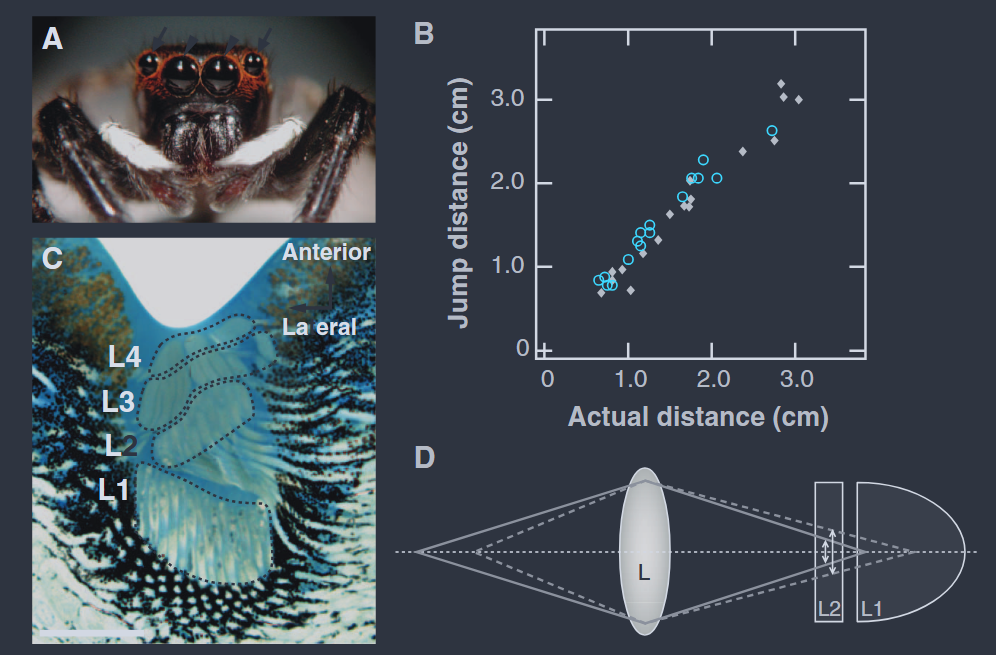
This mechanism was tested directly by exploiting chromatic aberration. Although the spider’s principal photoreceptors are tuned to green light, they retain weak sensitivity to longer wavelengths. As shown in Figure 7, the green-sensitive receptor responds to red light at roughly one-tenth the strength of green.
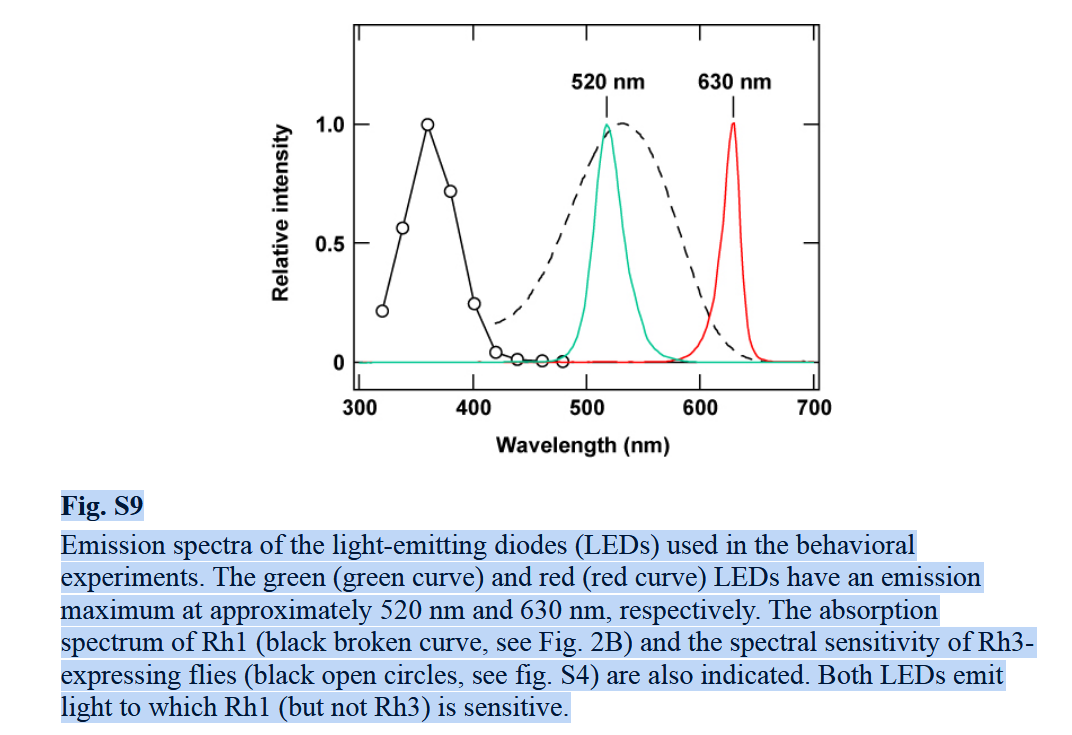
In the experiment, targets were therefore illuminated with red light whose intensity was increased about tenfold, so that the photoreceptors were activated as strongly as under green illumination. Brightness was thus matched at the receptor level. Because red light has a longer wavelength, however, the same amount of retinal defocus corresponds to a different object distance (Figure 8)
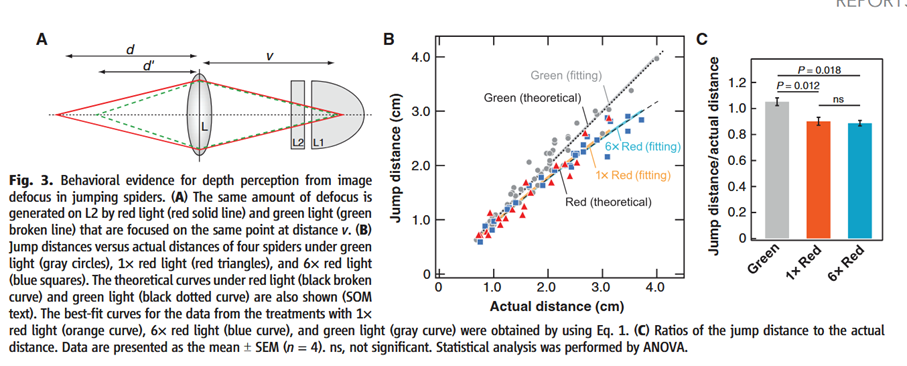
Under red illumination, spiders consistently underestimated distance and jumped short, even when red intensity was increased sixfold beyond this matching level, ruling out brightness as an explanation. These results confirm that jumping spiders infer distance from image defocus caused by chromatic aberration.
Portia does not merely see sharply. It sees details that should be impossible at its scale. Although this visual system requires longer sampling times, it provides information about who the prey is, what it is doing, and how it is oriented. With experience, Portia can recognize particular prey and anticipate how they are likely to respond. These perceptual capacities support the development of elaborate cognitive strategies, and in turn, it is these strategies that give evolutionary value to such perceptual refinement. In the next part, we will see Portia not only as an exceptional visual system, but as an exceptional strategist.
***
For more essay you can follow my monthly substack or directly catch up on my latest posts there.
Enjoying the content on 3QD? Help keep us going by donating now.